| 首页 > 新闻资讯 > 染色体结构捕捉技术 |
染色体结构捕捉技术自从人类开始使用显微镜观察细胞以来,细胞核内的丝状染色体构造一直为科学家所好奇。随着逐步建构的遗传学与生物化学概念,包括:由四个硷基构成密码的DNA是生物体内的主要遗传物质,影响性状表现的基因位在染色体上,DNA是由双股螺旋分子所构成,强化子影响启动子对其所属基因的表现调控等。  图一 3C技术原理与衍生之4C、5C及Hi-C技术。
他们的实验首先使用低浓度福马林处理细胞,使距离相近的丝状染色体产生键结,再用限制酶将大片段DNA切成小片段,使得键结的DNA序列形成X型构造(图一),接着用接合酶将X型DNA分子的两端进行分子内接合,以产生8字型或丝带状DNA构造。再以65℃左右高温进行反键结,将第一步产生的相邻DNA分子间键结打开,使接合的分子恢复成线状或环状。最后,以定量聚合酶连锁反应放大产物及后续定序。
由于相邻DNA序列有较高的机率于实验的第一步中产生键结,使得实验最后定量聚合酶连锁反应中,相邻序列有较多的反应产物。接着以定序结果反推序列于基因体上的位置,如此便能推测引子的目标序列与基因体内的其他特定序列,在空间上具有何种程度的交互作用,也就能依此建构一段特定DNA序列的染色体在细胞核内的3D结构。使用3C技术所得到的DNA结构图谱 ,可用来解释DNA结构上的改变,包括直链形成环形DNA,如何调控特定基因的表现。还有,由于DNA片段在空间上的相互作用,会造成染色体构型上的改变,进而影响相关基因的调控,因此可研究强化子和启动子如何影响目标基因的RNA转录。
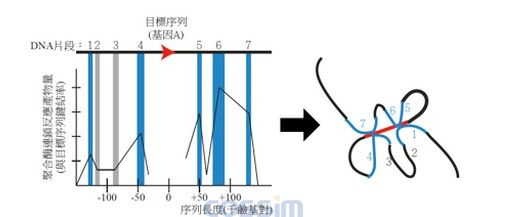 图二模拟3C技术所得之序列位置对照聚合酶反应产物图与DNA构型图谱。
左图模拟基因A经过3C技术所得之上下游序列片段(1到7)聚合酶连锁反应产量。由于聚合酶反应产物量与相邻DNA序列在实验过程中形成的键结率成正相关,因此可以由纵轴的数值反应出片段之间在空间上的远近,进而推测出长链DNA在空间上的相互作用关系图谱。序列片段1到7中,可能部分为已知的强化子,另外功能从未被描述但与基因A有相互作用的其他片段,则可能是具有功能的强化子。因此可以藉由3C技术得到新的基因调控片段与机制。另外,以往功能从未被描述的DNA序列,也可因为3C技术的应用,得以让研究人员从序列与基因片段的交互作用中,预测出新的基因表现调控机制。因此,3C技术的发展为基因表现的研究带来了相当大的突破。 然而3C技术也不是全然没有限制,由于大部分双倍体生物的基因只有一对序列,针对基因体中数百万硷基对进行放大时,稀少片段所得到的讯号往往容易被非专一性键结的杂讯给覆盖。因此对照组挑选、谨慎的实验设计、实验数据的正确解读,有可能会造成3C实验结果判定上的困难。另外,由于3C技术针对目标序列以聚合酶连锁反应进行放大,在一次实验设计中要探讨整个基因体序列的交互作用必须准备相当多的引子对,仍然限制全基因体3D结构的分析 。
随着生物晶片以及更便宜且高通量的次世代定序陆续发展,以3C为基础的4C、5C或 HiC 等技术,被设计并应用以解构一条带有数亿硷基对染色体的构型 ,或提升到数千个硷基对以下的解析度,使科学家能够更精确地预测可能的基因调控序列及染色体结
文章出自:科信仪器 转载时必须以链接形式注明作者和原始出处及本声明。
|
| 首页 > 新闻资讯 > 染色体结构捕捉技术 |







